Cognitie en neurowetenschappen
Inhoudsopgave
Hersenanatomie en werkingDe ontwikkeling van de hersenen
Geheugen, aandacht en bewustzijn
Ruiken
Zien
Horen
Elektrofysiologisch
Het brein in beeld gebracht
Hersenanatomie en werking
Structuur en functie van het neuron
Alle neuronen hebben grofweg dezelfde structuur:
- Dendrieten (ontvanger)
- Cellichaam / soma
- Axon (zender)
- Kan bestaan uit verschillende vertakkingen die "collaterals" genoemd worden
Signalen worden doorgegeven op basis van een actiepotentiaal, ofwel een "alles of niets" actie. Een neuron kan niet "half" vuren. De dendrieten ontvangen signalen en op het moment dat er genoeg signalen ontvangen worden, wordt er een actiepotentiaal afgevuurd. Hierover later meer.
Enkele interessante feiten over de hersenen:
- De hersenen bevatten ca. 86 miljard neuronen
- Elk neuron is verbonden met ongeveer 10.000 andere neuronen
- De hersenen maken ongeveer 2% uit van het totale lichaamsgewicht
- We verliezen ongeveer 1 corticaal neuron per seconde
- Tussen de 20 en 90 jaar verliezen we pakweg 10% van onze corticale neuronen
- Hersengroote is sterk genetisch bepaald, maar eeneiige tweelingen hebben geen identieke hersenen. (Nature vs nurture, in dit geval bepaalt nurture.)
- Mannen hebben grotere hersenen, maar vrouwen hebben meer vouwen en compenseren door een groter oppervlakte (surface area)
Een verbinding tussen neuronen is een synaps. Dit betekent dat je kunt spreken van een presynaptisch neuron en een postsynaptisch neuron. Respectievelijk voor de synaps en na de synaps.
Een geactiveerd neuron vuurt een actiepotentiaal af door het axon heen. Aan het einde van het axon worden vervolgens neurotransmitters losgelaten die op hun beurt invloed uitoefenen op de dendrieten van het postsynaptisch neuron door zich te binden aan specifieke receptoren voor die neutotransmitters (of direct aan het cellichaam). Hierdoor ontstaat een synaptisch potentiaal. Dit zorgt voor een passieve stroom door de dendrieten en het cellichaam van het postsynaptisch neuron loopt. Als al deze passieve stromen bij elkaar opgeteld een groot genoeg potentiaal opwekken, volgt er een actiepotentiaal die door het axon gevuurd wordt (bijvoorbeeld naar de volgende synaps). Het meten van al deze passieve stromen is de basis van EEG-scans.
De opbouw van synaptisch potentiaal gebeurt dus door passieve geleiding. Dit werkt alleen op korte afstanden, door de weerstand van de omliggende materie. Actieve geleiding zoals die van een actiepotentiaal, is daarentegen wél geschikt voor langeafstandssignalering tussen neuronen.
Let op: sommige neuronen signaleren puur elektrisch en niet door middel van neurotransmitters, zoals in de retinal gap junctions.
Elektrische signalering en de actiepotentiaal
Ieder neuron heeft een celmembraan. De binnenkant van het membraan is negatief geladen (ca. -70mV) ten opzichte van de buitenkant van het membraan, en bij het axon is deze voorzien van voltage-gated ionkanalen. Een actiepotentiaal verloopt als volgt:
- Passieve stroom loopt langs het membraan van het axon. Hierdoor gaan Na+-kanalen open.
- Na+ stroomt de cel in en zorgt voor depolarisatie
- Rond -50mV depolariseert de cel volledig in een alles-of-niets actie omdat Na+ steeds sneller de cel binnen stroomt
- De cel laat K+ ionen door het membraan om weer een negatieve lading te krijgen: de cel repolariseert
- Voor een kort moment ontstaat er hyperpolarisatie. Dat wil zeggen dat de binnenkant van het celmembraan nóg negatiever is, waardoor opnieuw activeren bijzonder lastig is. Daardoor kan het signaal niet achteruit bewegen.
- De cel herstelt zich en is weer klaar voor een nieuwe actiepotentiaal
Op deze manier loopt een signaal door het hele axon heen.
Als axonen gemyeliniseerd zijn (omhuld met myeline), dan gaat het signaal onder de myeline passief verder. Myeline is een witte, vetachtige substantie die in feite als bescherming en als isolator werkt, maar die ervoor zorgt dat signalen sneller en verder kunnen reizen. De actiepotentiaal wordt verder overgedragen tussen de zogenaamde "knopen van Ranvier". Dit zijn de stukjes axon waar geen myeline zit.
Er zijn verschillende aandoeningen waarbij de myeline kapot gaat. Een goed voorbeeld hiervan is MS (Multiple Sclerose).
Chemische signalering en het postsynaptische neuron
Uiteindelijk komt de actiepotentiaal terecht aan het einde van het axon: de axon terminal. Hier laat deze neurotransmitters in de synaptische spleet (synaptic cleft). Proteïnereceptoren op het postsynaptisch membraan binden zich aan de neurotransmitters. (Dit zijn de zogenaamde transmitter-gated ionkanalen.) Een lokale stroom van Na+, K+ of Cl- vindt plaats, afhankelijk van de neurotransmitter.
GABA is een voorbeeld van een inhibiterende neurotransmitter, omdat deze Cl- kanalen opent en dus zorgt voor een grotere polarisatie (en het dus lastiger maakt voor het neuron om de -50mV voor de actiepotentiaal te bereiken). De cel wordt van binnen negatiever en is dus moelijker te activeren. Glutamaat is daarentegen een voorbeeld van een exciterende neurotransmitter, die juist zorgt dat de actiepotentiaal makkelijker op te wekken is. GABA en glutamaat zijn de werkpaarden onder de neurotransmitters in de hersenen.
Belangrijk om op te merken, is dat het niet de neurotransmitters zijn die zelf de actiepotentiaal veroorzaken. De neurotransmitters werken in op de ionkanalen en de "status" van de ionkanalen bepaalt of er een actiepotentiaal komt.
Regulerende neurotransmitters zijn bijvoorbeeld:
- Serotonine
- Dopamine
- Acetylcholine
- Noradrenaline
De afgifte hiervan is vaak meer gelokaliseerd in de hersenen. D.w.z. in bepaalde hersengebieden. Dat wil echter niet zeggen dat de effecten ervan niet door de hele hersenen kunnen doorwerken, want ze bepalen toch de signalering.
De amplitude van een actiepotentiaal is altijd hetzelfde. Het is immers een alles-of-niets actie. Wat wél verandert, is de spike rate ofwel hoe vaak een neuron vuurt (preciezer: het aantal actiepotentialen per seconde). Neuronen die vaak vuren in bepaalde situaties, zijn vaak omringd door andere neuronen die vaak vuren in dezelfde omstandigheden. Dit vormt de basis voor de lokalisatie van functie in de hersenen. Het is een logisch gevolg van hoe een neuraal netwerk functioneert. Dat betekent dus ook dat de functie van bepaalde hersengebieden sterk afhankelijk is van de input die dat deel van de hersenen krijgt. Zonder input geen output en zonder output is er geen functie vervuld.
Een gedachtenexperiment: stel dat de auditieve signalen naar de visuele hersengebieden worden gestuurd. Door de verandering van de input verandert automatisch de functie van dit gebied. Lokalisatie van functie is daarom een enigszins discutabel onderwerp.
De grove organisatie van de hersenen
De hersenen bestaan onder andere uit grijze stof, witte stof en cerebrospinale vloeistof.
- De grijze stof bestaat met name uit cellichamen van neuronen
- De witte stof bestaat vooral uit axonen en gliacellen (steuncellen)
- De cerebrospinale vloeistof vervoert afvalstoffen, vervoert signalen en biedt een soort stootkussen voor de hersenen.
De "gevouwen" buitenste laag van de hersenen (de cerebrale cortex) bestaat uit grijze stof. Daaronder ligt de witte stof en daaronder, nog verder in de hersenen, zijn er weer andere structuren in grijze stof (de subcortex), zoals de basale ganglia, het limisch systeem en de diencephalon (de tussenhersenen).
Er zijn verschillende tracten (tracts), een soort zenuwverbindingskanalen:
- Associatietracten (association tract)
- De verbindingen in de witte stof (immers vooral axonen) die binnen dezelfde hemisfeer blijven
- Projectietracten (projection tracts)
- Verbindingen tussen corticale structuren en subcorticale structuren (zoals hersenstam en ruggenmerg)
- Commissurale tracten (commissural tracts)
- Verbindingen tussen regio's in verschillende hemisferen. Het corpus callosum is hier een goed voorbeeld van.
Refereren naar onderdelen van de hersenen gaat op de volgende manier:
dorsal
superioranterior
rostralposterior
caudalventral
inferiorDaarnaast spreek je van medial wanneer je richting het centrum van een structuur gaat (zoals de hele hersenen, maar ook een bepaalde regio binnen de hersenen) en lateral wanneer het richting de buitenkant van de hersenen is.
Er zijn vier verschillende secties (doorsnedes) die kunnen worden gemaakt:
- Coroaal (coronal)
- Verticaal, door beide hemisferen
- Saggitaal (saggital)
- Verticaal, door één hemisfeer
- Middenlijn (midline)
- Verticaal, tussen de hemisferen door
- Axiaal (axial)
- Horizontaal (per definitie door beide hemisferen)
De saggitale sectie loopt dus parallel aan de middenlijn, maar gaat slechts door één hemisfeer.
De cerebrale cortex
De cerebrale cortex bestaat uit twee "vellen" van grijze stof, die georgniseerd zijn als twee losse hemisferen. Door de evolutie heen zijn deze vellen steeds verder gevouwen, wat als voordeel biedt dat de totale oppervlakte ervan vergroot wordt. De bobbels heten gyri en de inkepingen heten sulci.
De cerebrale cortex is ca. 3mm dik en bestaat op de meeste plekken uit 6 verschillende lagen (met verschillende celtypes) die we de neocortex noemen (nieuwe cortex). Andere corticale regio's zijn de mesocortex (o.a. singulate gyrus en de insula) en de allocortex (o.a. primaire olfactoire cortex en hippocampus).
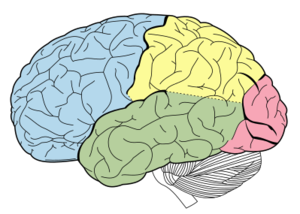
Het laterale gebied van de cerebrale cortex is opgedeeld in 4 kwabben: frontaal, pariëtaal, temporaal en occipitaal. Soms is de scheidingslijn hiertussen heel duidelijk, maar vaak ook niet. Tussen de frontale en temporale kwab is het verschil echter goed te zien en deze scheidingslijn heet de sylvian fissure of lateral fissure.
Er zijn vier manieren om hersengebieden te groeperen:
- Op basis van het patroon in gyri en sulci. Hoewel de vorm en grootte per individu kan verschillen, is het patroon altijd hetzelfde.
- Op basis van cytoarchitectuur, zoals Brodmann's area's. Dit waren oorspronkelijk 52 regio's, spiraal genummerd van binnen naar buiten (laag naar hoog).
- Op basis van functie. Wordt eigenlijk alleen gebruikt voor primaire motorische en visuele gebieden. Hogere corticale gebieden zijn lastig in te richten naar functie.
- Op basis van connectiviteit. Iedere regio heeft een ander connectiviteitsprofiel en met behulp van MRI kunnen regio's zo gesegmenteerd worden.
De subcortex
Onder de (lateraal gelegen) cerebrale cortex, is er een laag met witte stof (vooral gemyeliniseerde axonen) met daaronder de subcortex.
Basale ganglia
De basale ganglia zijn grote ronde massa's die in beide hemisferen te vinden zijn. Ze hangen "over" de thalamus heen in het midden van de hersenen. Ze zijn verantwoordeijk voor het reguleren van motorische activiteit en de programmering van start- en einde van actie. Daarnaast zijn ze gerelateerd aan het reward-system en het vormen van gewoonten en het leren van nieuwe vaardigheden.De meest bekende ziekten die te maken hebben met de basale ganglia zijn onder te verdelen in twee categorieën:
- Hypokinetische stoornissen
Te weinig beweging, zoals Huntington's- Hyperkinetische stoornissen
Overvloed aan beweging zoals Parkinson'sLibisch systeem
Het limbisch systeem is een regio van de subcortex die belangrijk is in het monitoren van en handelen naar de omgeving van het organisme (zowel nu als in het verleden). Het is van belang bij het detecteren én uitdrukken van emotionele uitdrukkingen.
- De amygdala is bijvoorbeeld verantwoordelijk voor het detecteren van angstwekkende en bedreigende stimuli.
- Delen van de cingulate gyrus detecteren emotionele en cognitieve conflicten.
- De hippocampus is van belang voor leren en geheugen.
- De mammilaire lichamen zijn belangrijk voor geheugen
- De olfactory bulbs zijn verantwoordelijk voor het reukvermogen en het feit dat ze direct ingeplugd zijn in het limbische systeem geeft aan hoe belangrijk geur is in het opwekken van emotie, gemoedstoestand en herinneringen
Diencephalon
Er zijn twee structuren die samen de diencephalon opmaken: de thalamus en de hypothalamus. De thalamus is in feite het station waar alle sensorische signalen langs gaan voordat ze naar de cortex gaan, met uitzondering van geur (die gaat immers via de olfactory bulbs rechtstreeks het limbisch systeem in). De thalamus zijn twee eivormige structuren die in beide hemisferen te vinden zijn. Vanuit de thalamus lopen projectietracten naar bijna alle regio's in de cortex en basale ganglia. Belangrijke namen zijn de lateral geniculate nucleus, die verantwoordelijk is voor het doorsturen van visuele signalen en de medial geniculate nucleus, die verantwoordelijk is voor het doorsturen van auditieve signalen.Het middenbrein en hindbrain (rhombencephalon)
Het middenbrein (mesencephalon) en de hindbrain (rhombencephalon) bestaat uit een groot aantal structuren. Belangrijke structuren zijn de superior colliculi, de inferior colliculi, het cerebellum, de pons en de medulla oblongata.
- Superior colliculi
- Onderdeel van een subcorticaal pad voor sensorische informatie. Betrokken bij programmering van snelle oogbewegingen.
- Inferior colliculi
- Onderdeel van een subcorticaal pad voor auditieve informatie.
- Cerebellum
- Kleine hersenen, verantwoordelijk voor behendigheid en accuratesse in beweging
- Pons
- De link tussen het cerebellum en het cerebrum. Ontvangt informatie om oog en lichaam te bewegen.
- Medulla oblongata
- Komt uit de pons voort en gaat over in het ruggenmerg. Reguleert vitale functies zoals ademen, slikken, hartslag en de slaap-waak-cyclus (circadiaans ritme).
Reacties
Er zijn nog geen reacties.Meld je aan met LinkedIn om te reageren